


Одной из центральных проблем современной эстетической медицины является лечение целлюлита. Это объясняется, с одной стороны, необходимостью удовлетворить запросы клиентов, желающих избавиться от данного эстетического недостатка, с другой – изучить закономерности его возникновения и возможности эффективной коррекции.

Елена Дарбанова
Руководитель компании «Диона»
Les nouvelles esthetiques, 3/2014
О причинах развития целлюлита существует много гипотез, которые порой противоречат друг другу. Некоторые исследователи связывают этот процесс с изменениями соединительной ткани, его основным диагностическим признаком считают «апельсиновую корку», а стадии развития — от 1 до 4 — определяют степенью выраженности данного симптома.
На наш взгляд, изменения кожи на тканевом уровне являются вторичным фактором развития процесса, в то время как причина возникновения целлюлита связана с нарушением работы клетки. Сам термин «целлюлит» происходит от латинского слова cellula — клетка, а суффикс -ит, присоединяемый к латинскому или греческому слову, обозначает воспалительный характер патологии. Таким образом, термин «целлюлит» в его точном этимологическом смысле означает «заболевание, вызванное воспалением клетки», что отражает истинную природу его возникновения.
Среди многочисленных публикаций на эту тему нет материалов, касающихся изменения клеток при целлюлите. Многие специалисты считают целлюлит только косметическим недостатком, однако мы полагаем, что это — серьезное заболевание, связанное с изменением жировой клетки, энтропия которой ведет к формированию целого ряда нарушений в соединительной ткани.
Поэтому понимание механизма работы адипоцита в нормальном и «целлюлитном» режиме сможет приблизить специалистов к объяснению процесса формирования данной патологии.
ФИЗИОЛОГИЯ ЖИРОВОЙ ТКАНИ
Известно, что жировая ткань, состоящая из клеток — адипоцитов, выполняет наряду с другими важными функциями роль своеобразного энергохранилища, в котором находится «стратегический запас» организма в виде триглицеридов (ТАГ). При их расщеплении образуются моноглицериды (МАГ), свободные жирные кислоты и молекулы АТФ, благодаря которым сохраняется высокий энергетический потенциал клетки.
На мембранах жировых клеток расположены молекулярные структуры — альфа- и бета-рецепторы, активация которых запускает процессы липогенеза и липолиза.
Эти процессы имеют различные, но дополняющие друг друга механизмы. Альфа-2-рецепторы ответственны за фазу липогенеза — образование и запасание жиров, а бета-рецепторы — за липолиз, т.е. фазу распада жиров. Рецепторы функционируют под контролем гормонов: липогенез регулируется инсулином, который стимулирует выработку специальных ферментов, обеспечивающих синтез жира; на фазу липолиза оказывают влияние гормоны прямого действия (адреналин, норадреналин) и гормоны косвенного действия (лептин). Они регулируют активность липазы жировой ткани, часто называемую «гормоночувствительной липазой». Следовательно, физиологический сбой организма, связанный с его гормональной перестройкой, может привести к нарушению механизма запасания и мобилизации жира в адипоцитах.
Гормональные изменения и состояние жировой клетки
Пубертат и климакс, вызывающие гормональную перестройку организма, становятся тем опасным периодом для женщины, когда жировая клетка может подвергнуться патологическим изменениям. Слабым звеном являются области ягодиц, бедер и частично живота, поскольку именно в этой части жировой ткани на адипоцитах преобладают альфа-2-рецепторы, и, следовательно, фаза запасания жира становится для них ключевой. Для реализации этой фазы необходимо наличие свободных жирных кислот и глюкозы, которые проникают через мембрану адипоцита внутрь клетки, где и происходит синтез жиров в виде триглицеридов. По мере накопления образующихся жиров адипоцит увеличивается в размере.Но до тех пор, пока плазматическая мембрана клетки осуществляет, с одной стороны, жесткую барьерную функцию, а с другой — избирательно пропускает в цитозоль необходимые вещества, адипоцит находится в увеличенном, но физиологичном состоянии.
БИОЛОГИЧЕСКАЯ МЕМБРАНА
Можно предположить, что лишь изменения функциональной активности биологической мембраны приводят к сбою работы самой клетки, поскольку мембрана, являясь границей между внеклеточным матриксом и внутриклеточной средой, определяет множество биохимических процессов во время их взаимодействия. Это касается, прежде всего, стабилизации электрохимического, водного, ионного и энергетического баланса. Избирательная проницаемость мембраны необходима для поддержания разницы концентраций ионов натрия, калия, кальция, магния, хлора во внеклеточной среде и внутри клетки, которая может отличаться в тысячи раз (рис.1).

Рис.1 Различия в составе ионов внутри клетки и во внеклеточной среде
Известно, что все клеточные мембраны построены в основном из липидов, белков и углеводов. Структурной основой мембраны является бимолекулярный слой, образование которого обусловлено особенностями взаимодействия с водой мембранных липидов, среди которых преобладают фосфолипиды.
Условно в молекуле фосфолипида выделяют три части — гидрофильную головку, тело и гидрофобные хвосты. Последние, соединяясь друг с другом, выпускают гидрофильные головки наружу. Так образуется бимолекулярный фосфолипидный слой (бислой), который и служит каркасом биологической мембраны. Кроме того, в мембране присутствует холестерин (холестерол), который является основой для клеточных мембран. Располагаясь между фосфолипидными молекулами, он упорядочивает бислой в жидком состоянии и «разупорядочивает» в гелеобразном, участвуя, таким образом, в цикличном процессе фазового перехода мембраны из жидкокристаллического состояния в гелеобразное.
В покое мембрана поляризована: цитоплазма имеет отрицательный заряд, а внеклеточная среда — положительный. При возбуждении происходит деполяризация мембраны, потенциал внутри клетки становится положительным; после снятия напряжения восстанавливается поляризация.
В процессе фазового перехода в липидном бислое образуются сквозные каналы радиусом
- интегральные — пронизывают плазмалемму насквозь;
- полуинтегральные — занимают лишь часть мембраны;
- поверхностные, или периферические — предназначены избирательно транспортировать вещества из внеклеточного матрикса в цитозоль и обратно, поддерживая, таким образом, необходимую для гомеостаза концентрацию веществ по разные стороны мембраны.
Трансмембранный транспорт
Существует 4 механизма трансмембранного транспорта.
- Ионные каналы — Na+, К+, Са2+, Cl — осуществляют селективный перенос ионов по электрохимическому градиенту, т.е. из области большей концентрации вещества в меньшую, без затрат энергии, производя так называемый «пассивный транспорт».
- Ионные насосы — Na+, К+, Са2+ — транспортируют специфические вещества против электрохимического градиента, т.е. осуществляют «активный транспорт», на выполнение которого требуются затраты энергии молекул АТФ, поскольку выкачивание происходит из области меньшей концентрации веществ в большую. Эту функцию выполняют особые ферменты, специфичные для каждого вида переносимых субстанций — Na+ — АТФаза, K+ — АТФаза, Са2+ — АТФаза, т.е. для каждого вида насоса имеется специальный фермент. Соединение фермента с переносимыми ионами меняет конформацию активного центра АТФазы, что, в свою очередь, активизирует ее способность гидролизовать молекулы АТФ.
- Транспортеры ионов — это системы ионного симпорта, такие как Na+ — глюкоза, Na+ — K+ — Cl—, Na+ — фосфат, и ионного антипорта — Na+ — Са2+ — обменник и т.д. Перенос веществ осуществляется как по электрохимическому градиенту, так и против него, с использованием энергии гидролиза АТФ.
- Водные каналы, или аквапорины («водная пора»), обеспечивают быстрое прохождение воды внутрь клетки. Они представляют собой интегральные белки, состоящие из нескольких полипептидных цепей, которые, сворачиваясь в спиралеобразную форму, открывают пору, проницаемую для воды (рис. 2). Последние исследования, отмеченные Нобелевской премией, показали, что многие из аквапоринов могут строго избирательно транспортировать определенные вещества как внутрь клетки, так и вовне. Для этой цели у аквапоринов существуют специальные, положительно заряженные фильтры, пропускающие только молекулы воды и того вещества, за транспорт которого отвечает отдельный, специальный канал.
На сегодняшний день открыты 13 видов аквапоринов, особых для каждого типа клеток, так как, несмотря на наличие универсальной структуры в строении, существуют и специфические особенности, которые определены их функциональной природой.

Рис.2. Аквапорины (AQP)

Рис.3 Роль аквапоринов AQP7 в транспорте глицерина через мембрану адипоцитов.
Одной из задач жировых клеток является депонирование ТАГ (триацилглицеринов), предназначенных для гидролиза АТФ. Реагируя на сигналы инсулина, адипоцит с помощью симпорта натрий — глюкоза вводит глюкозу в цитозоль, затем ее поглощает, синтезирует и депонирует ТАГ. Эту фазу мобилизации жира сменяет фаза расщепления, гидролиза ТАГ с образованием глицерина (МАГ) и свободных жирных кислот. Глицерин (МАГ) через аквапорин AQP7 выводится из клетки (рис. 3) и, будучи веществом гидрофильным, растворяется в плазме крови.
Изменение баланса между липолизом и липогенезом в пользу последнего ведет к увеличению размера адипоцита, а уменьшение размера жировой клетки происходит при преобладании фазы липолиза. Так функционирует жировая клетка в нормальном режиме.
НАРУШЕНИЕ РАБОТЫ БИОЛОГИЧЕСКОЙ МЕМБРАНЫ И ВОЗМОЖНЫЕ ПОСЛЕДСТВИЯ
Известно, что плазматическая мембрана чувствительна к физиологическим сдвигам в организме, в зависимости от которых она меняет свою проницаемость для отдельных веществ. Избыточное поступление насыщенных жирных кислот нарушает структуру бислоя плазмалеммы, так как содержание в мембране холестерина значительно увеличивается, баланс фосфолипидов и холестерина изменяется. Все это приводит к увеличению микровязкости липидного слоя мембран и влечет за собой нарушение фазового перехода. Это, в свою очередь, снижает проницаемость бислоя для транспорта веществ по обе стороны плазмалеммы, поскольку меняется конформация белков и происходит ухудшение работы ферментативных систем. Накопление в мембране холестерина ингибирует Na+, K+ — АТФазу и Са2+ — АТФазу, которые являются одними из ключевых ферментов гомеостаза клетки. Скорость его распада уменьшается с ростом микровязкости липидов, в результате нарушается ферментативная реакция, и насос, выкачивающий из клетки Са2+, превращается в канал, т.е. ионы кальция начинают поступать по электрохимическому градиенту концентрации. В результате этого происходит повышение содержания кальция в цитозоле.
О последствиях этого процесса свидетельствуют исследования биофизиков, доказавших, что повышение внутриклеточной концентрации Са2+ может индуцировать отвердение липидного монослоя, обращенного к цитозолю.
Таким образом, изменения гормонального статуса организма и/или повышенное потребление углеводов и насыщенных твердых жиров приводят к выработке печенью излишнего холестерина, который транспортируется по кровеносному руслу липопротеидами очень низкой плотности (ЛОНП), встраивается в мембрану клетки, тем самым увеличивается микровязкость липидного слоя. Это ведет к изменению конформации белков, уменьшению активности ферментативных систем, Са2+ — АТФазы, отвечающих за вывод кальция из клетки. Накопление в клетке избыточного кальция приводит к дальнейшему отвердению, т.е. обызвествлению внутреннего липидного слоя. Фазовый переход из гелевого состояния в жидкокристаллическое нарушается, поры не открываются для прохода, и мембрана становится непроницаемой для транспортировки веществ как во внутрь клетки, так и вовне.
Продукты гидролиза триглицеридов — глицерин, выводящийся из жировой клетки через аквапорин AQP7, жирные кислоты, а также молекулы АТФ — остаются во внутриклеточном пространстве, где, по всей видимости, опять синтезируются в ТАГ. В то же время ионы натрия, калия, хлора и другие вещества оседают внутри клетки. В связи с изменением концентрации веществ по обе стороны плазматической мембраны нарушается взаимодействие цитозоля с внеклеточным пространством. Изменение количества ионов натрия, хлора, кальция в клетке ведет к уменьшению необходимой их концентрации уже в соединительной ткани. Это приводит к нарушению электропотенциала, изменению водного баланса и, соответственно, молекулярного состава межклеточного матрикса. Таким образом, наступает перицеллюлярная воспалительная реакция в соединительной ткани. В ответ на это фибробласты начинают усиленно продуцировать коллаген и эластин, производя их избыточное количество. Утилизация металлопротоиназами (ММР) излишнего коллагена замедляется, так как они относятся к кальцийзависимым протеиназам.
Необходимое количество внеклеточного кальция, которого в нормальном режиме работы клетки должно быть почти в 10 000 раз больше, чем внутриклеточного, вследствие ингибирования мембраны уменьшается. Это вызывает изменение активности ферментативных реакций под воздействием ММР. Избыточный коллаген и эластин укладываются неупорядоченными пластами. Образовавшаяся фиброзная ткань начинает оказывать компрессионное давление на капилляры.
Изменение водного баланса ведет к возникновению отеков. Все это отражается на внешнем виде кожи как проекции внутренних изменений клетки, достигших стадии целлюлита.
Таким образом, можно с большой долей вероятности сказать, что процесс образования целлюлита связан с цепочкой патологических изменений, происходящих в следующей последовательности.
Гиперхолестеризация плазмалеммы, увеличение микровязкости липидного слоя, нарушение фазового перехода от жидкокристаллического состояния мембраны в гелевое
![]()
Изменение работы всех структур межмембранного транспорта веществ, ингибирование ключевых ферментов ионных насосов, в частности Са2+ — АТФазы
![]()
Инициация процесса превращения насоса, откачивающего избыток кальция, в канал, который действует противоположно
![]()
Поступление из матрикса в цитозоль по электрохимическому градиенту большого количества Са2+, который оседает на внутреннем монослое мембраны, приводя к его обызвествлению
![]()
Клетка становится непроницаемой для всех веществ как на вход, так и на выход. Воспаление клетки приводит к перецеллюлярному фиброзу соединительной ткани со всеми вытекающими из этого клиническими последствиями
СОБСТВЕННЫЕ ИССЛЕДОВАНИЯ
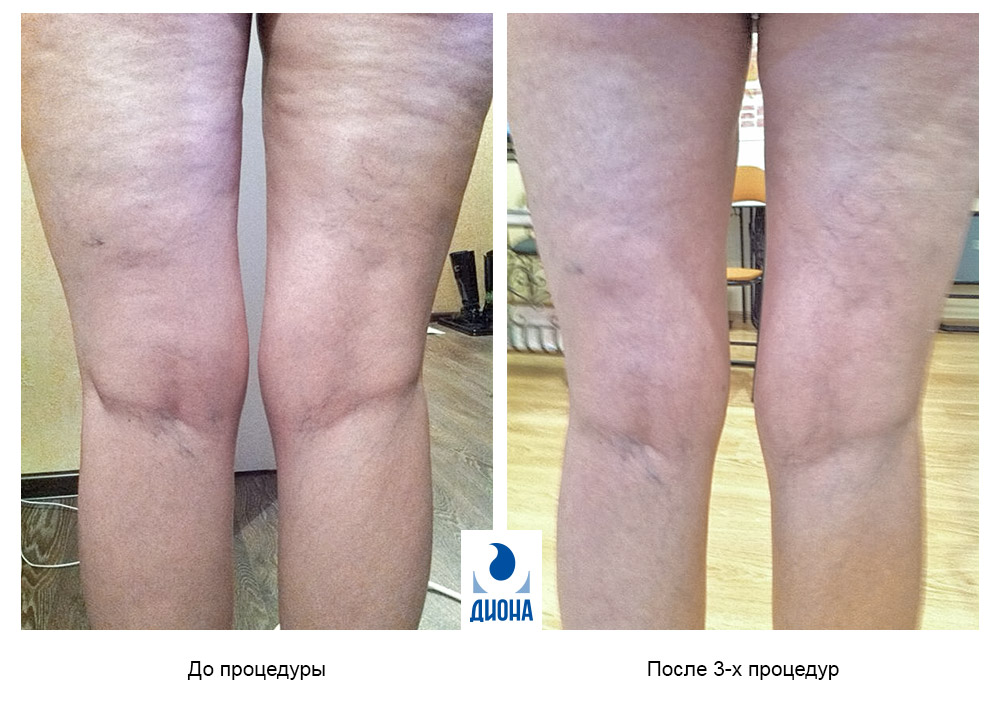
Фото 1. Пациентка А. (51 год) до и после проведения 3-х процедур.
На поиск понимания природы целлюлита нас натолкнули собственные результаты, которые были получены в ходе практического применения метода радиочастотного воздействия. Ранее нами было показано, что при применении RF-технологий, в частности при использовании излучения в лечебной дозе с частотой 6 МГц, выраженный оздоравливающий результат отмечается не за счет индукции тепла, а благодаря созданию осцилляторного эффекта, т.е. воздействию электромагнитного поля высокой частоты. Изменение соединительной ткани было подтверждено исследованием, проведенным в Испытательном лабораторном центре ЦКБ РАН. Было выявлено повышение показателя эластометрии на 27% при отсутствии побочных эффектов.
Данные, подтверждающие изменение структуры кожи при использовании RF-технологий, явились основанием для применения этого аппарата при лечении целлюлита. Действительно, эффект от процедур был впечатляющим (фото 1): уже после трех сеансов наблюдался выраженный результат.
Теоретическое обоснование
Этот факт стал отправной точкой для про ведения теоретических исследований. Систематизация достижений смежных наук позволила нам приблизиться к пониманию процесса образования целлюлита, связанного с патологическим изменением жировой клетки, влекущее, в свою очередь, локальное изменение соединительной ткани.
В наших предыдущих исследованиях отмечалось, что при воздействии переменным электромагнитным полем высокой частоты в организме происходят определенные биохимические изменения. Их характер определяется способностью высокочастотного электромагнитного излучения взаимодействовать с заряженными частицами биологических тканей — ионами, белками, метаболитами, нуклеиновыми кислотами, полярными головками фосфолипидов, а также молекулами воды.
Усиленные колебания водородных связей диполя воды при воздействии излучением с частотой 6 МГц (6 млн раз в секунду) изменяет ее конформацию. Образуются гексагональные структуры молекул воды, что ведет к гидратации коллагена и, таким образом, увеличению уровня связанной воды. Восстанавливается устойчивая структура тропоколлагена, так как за счет водно-мостиковой поддержки укрепляются поперечные связи коллагеновых фибрилл.
В свою очередь, под действием электромагнитной волны высокой частоты происходит конформационное изменение и молекул коллагена. Выстраиваясь по силовым полям в правозакрученную альфа-спираль, тропоколлаген возвращается к своему нативному состоянию. Это и приводит к высокому лечебному результату, где омоложение кожи является хоть и побочным, но очень впечатляющим результатом. Лечебный эффект, возникающий при терапии целлюлита высокочастотным электромагнитным полем, можно также объяснить изменениями на клеточном уровне.
Мишенями резонансного колебания электромагнитного поля являются мембранные образования клеток и внутриклеточных органелл, в которых под действием излучения происходят структурно-функциональные изменения. В результате создается физико-химическая основа для коррекции процессов метаболизма, связанного с переносом протонов и электронов в клеточных мембранах, а уже на этой основе возникают последовательные неспецифические реакции клетки и организма в целом.
Справедливость этого тезиса была подтверждена большим количеством экспериментов, в ходе которых исследовали изменения конформационных состояний белковых молекул, в том числе и белков клеточных мембран. Модели взаимодействия электромагнитного излучения с белковыми молекулами показывают возможность синхронизации внутренних колебаний некоторых цепей, и за счет этого — концентрации энергии, достаточной для изменения конформации в макромолекулах. Так, при воздействии электромагнитного поля происходит изменение конформации белков плазмы крови, которые инициируют активизацию метаболических реакций. Изменение конформации белков крови, по всей видимости, приводит к функциональной активности апобелков, в частности АпоА1, являющихся центральным звеном в действии фермента — лецитинхолестеринацетилтрансферазы (ЛХАТ), которая катализирует липопротеиды высокой плотности (ЛПВП), обеспечивающие выведение избыточного холестерина из клеток тканей в печень. Под действием ЛПВП соотношение холестерина и фосфолипидов плазмалеммы меняется в сторону физиологической нормы. Это ведет к восстановлению цикличности фазового перехода мембраны из гелевого состояния в жидкокристаллическое. Экспериментальные подтверждения существования конформационных переходов под действием электромагнитного поля дают нам возможность предполагать, что аквапорины, будучи интегральными белками, при действии лечебной дозы электромагнитного излучения могут менять свою конформацию и, скручиваясь по силовым полям спиралеобразно, восстанавливают функцию водного канала, что, в свою очередь, активизируют метаболические реакции клетки.
Вода, как уже было сказано ранее, является первичной мишенью воздействия электромагнитного излучения, под влиянием которого происходит структуризация ее молекулярных связей. Измененная под действием электромагнитного излучения вода проникает через водные каналы в цитозоль.
Это ведет к восстановлению водного баланса и, следовательно, к усилению метаболизма внутри клетки.
Еще одним доказанным эффектом воздействия ЭМП является повышение активности целого ряда ферментов, относящихся к классу гидрогеназ: Na+/K+ — АТФаза, Ca2+ — АТФаза. Это дает возможность предположить, что повышение функциональной активности Са2+ — АТФазы на фоне общего процесса нормализации работы клетки приводит к восстановлению работы насоса, откачивающего избыточный кальций из внутриклеточного пространства.
Таким образом, цепочка реакций под действием тока высокой частоты способствует нормализации уровня холестерина, что инициирует восстановление фазового перехода, во время которого происходит открытие пор для прохождения веществ как внутрь клетки, так и вовне ее. Гидратация цитозоля, физиологическое функционирование кальциевого насоса и других ферментативных систем приводят к вымыванию кальция, осевшего на внутреннем монослое мембраны. Восстановленные функции аквапоринов позволяют нормализовать все трансмембранные механизмы по переносу веществ из цитозоля во внеклеточную среду. Так приходят в физиологичное равновесие метаболические процессы, связанные с поддержанием необходимой концентрации веществ по обе стороны мембраны. Конформационные изменения молекулярных структур транспортных интегральных белков активизируют процессы восстановления метаболического баланса внутриклеточного и внеклеточного гомеостаза. В результате жировая клетка возвращается к своему физиологическому состоянию.
Благодаря большому количеству экспериментальных исследований в области биофизики, биохимии, а также собственному практическому опыту, мы можем с большой долей вероятности заключить, что радиочастотное воздействие приводит к выраженному оздоравливающему результату как на тканевом, так и на клеточном уровне. Однако всегда надо помнить, что биологические эффекты от электромагнитного излучения полностью зависят от параметров воздействующих полей. Следовательно, понятие «лечебная доза» становится ключевым при выборе метода лечения целлюлита радиочастотным током. Ее характеристики определяются техническими параметрами аппарата и составляют 50 Вт входной мощности и 6 МГц частоты.
Рекомендуемая литература
- Baturina G.S., Katkova L.E., Ilyaskin A.V., Solenov E.I. Regulatory volume decrease. Role of water channels. The seventh international conference on bioinformatics of genome regulation and structure/systems biology. Novosibirsk,
20–27 June 2010, pp. 39. - Брежестовский П.Д. «Архитектура рецепторуправляемых ионных каналов биологических мембран». «Биофизика», 2011, т. 56, вып.1, с.
62–73. - Васильева Е.Г. «Механизм влияния электромагнитных полей на живые организмы». «Вестник АГТУ», 2008, № 3 (44), с.
186–190. - Владимиров Ю.А. «Кальциевые насосы живой клетки». Соросовский образовательный журнал, 1998, № 3, с.
24–26. - Гапеев А.Б., Романова Н.А., Чемерис Н.К. «Структурные изменения хроматина лимфоидных клеток под воздействием низкоинтенсивного электромагнитного излучения крайне высоких частот на фоне воспалительного процесса». «Биофизика», 2011, т. 56, вып. 4, с.
688–695. - Дарбанова Е.М. «Механизм действия радиочастотного лифтинга на биологические ткани». Les Nouvelles Esthetiques, 2013, №3, с.
84–90. - Иляскин А.В. «Экспериментально-теоретическое исследование водно-электролитного обмена клетки транспортного эпителия». Автореферат диссертации, Новосибирск, 2013, с.
2–12. - Калюжная Л.В. «Воздействие физических факторов на плазму крови». Тезисы VII Всесоюзного семинара «Использование низкоинтенсивного электромагнитного излучения в биологии и медицине», Москва, 1989, с. 54.
- Кругликов И.Л. «Противоречия эстетической медицины. Загадка целлюлита». «Эстетическая медицина», 2013, № 4.
- Малюкин Ю.В., Кавок Н.С., Боровой И.А., Степаненко А.М., Малюкина М.Ю., Петрухин П.А. «Исследование влияния адреналина на трансмембранный потенциал плазматической мембраны гепатоцитов крыс разного возраста с помощью флуоресцентных зондов». «Биофизика», 2011, т. 56, вып. 3, с.
475–480. - Мосин О.В. «Аппараты магнитной обработки воды». «Сознание и физическая реальность», 2011, т. 16, № 9, с.
16–32. - Панин Л.Е., Мокрушников П.В. «Воздействие андрогенов на активность Na+, K+ — АТФазы эритроцитарных мембран». «Биофизика», 2014, т. 59, вып.1, с.
127–133. - Пашук Л.К. «Биологическая мембрана и гомеостаз». Les Nouvelles Esthetiques, 2010, № 2.
- Перельмутер В.М., Ча В.А., Чуприкова Е.М. «Медико-биологические аспекты взаимодействия электромагнитных волн с организмом». Томский политехнический университет, 2007, с.
19–23. - Соловьева Н.И. «Основные металлопротеиназы соединительно-тканного матрикса». «Биоорганическая химия», 1994, т. 20, № 2, с.
143–152. - Тамашевский А.В., Козлова Н.М., Гончарова Н.В., Зубрицкая Г.П., Слобожанина Е.И. «Влияние холестерина на функциональную активность белков, ответственных за устойчивость лимфоцитов человека к действию ксенобиотиков». «Биофизика», 2011, т. 56, вып. 3, с.
455–464. - Титов В.Н. «Нарушение транспорта в клетки насыщенных жирных кислот в патогенезе эссенциальной гипертонии». «Клиническая лабораторная диагностика», 1999 , № 2, с. 5.
- Харакоз Д.П. «О возможной физиологической роли фазового перехода „жидкое — твердое“ в биологических мембранах». «Успехи биологической химии», 2001, т. 41, с. 337.
- Экспертное заключение по результатам испытаний эффективности применения аппарата Toplaser RF Esthetics Device RFV1. ЦКБ РАН, № 10135/591 от 27.09.2012 г.